Scientists discovered the part of the brain responsible for…
In my last post, I discuss one alternative to the modular theory of the mind/brain relationship: connectionism. Such a model is antithetical to modularity in that there are only distributed networks of neurons in the brain, not special-purpose processors.
One strength of the modular approach, however, is that it maps quite well to our folk psychology. And, much of the popular discourse surrounding research in neuroscience involves the celebrated “discovery” of the part of the brain responsible for X. A major theme of the previous posts is that the social sciences should be skeptical of the baggage of our folk psychology. But, is there not some truth to the idea that certain regions of the brain are regularly implicated in certain cognitive processes?
The earliest attempts at localization relied on an association between some diagnosed syndrome—such aphasia discussed in the previous posts—and abnormalities of the brain’s structure (i.e. lesions) identified in post-mortem examinations. For example, Paul Broca, discussed in my previous post, noticed lesions on a particular part of the brain in patients with difficulty producing speech. This part of the brain became known as Broca’s area, but researchers only have a loose consensus as to the boundaries of the area (Lindenberg, Fangerau, and Seitz 2007).
Furthermore, the relationship between lesions in this area and aphasia is partial at best. A century later, Nina Dronkers, the Director of the Center for Aphasia and Related Disorders, states (2000:60):
After several years of collecting data on chronic aphasic patients, we find that only 85% of patients with chronic Broca’s aphasia have lesions in Broca’s area, and only 50–60% of patients with lesions in Broca’s area have a persisting Broca’s aphasia.
More difficult for the modularity thesis, those with damage to Broca’s area and who also have Broca’s aphasia usually have other syndromes. This implies that the area is multi-purpose, and thus not a single-purpose language production module (see this book-length discussion Grodzinsky and Amunts 2006). One reason I focus on Broca’s area (apart from my interest in linguistics) is that it is considered the exemplary case for the modular theory quite dominant (if implicit) in much neuroscientific research (Viola and Zanin 2017).
Part of the difficulty with assessing even weak modularity hypotheses, however, is that neuroanatomical research continues to revise the “parcellation” of the brain. The first such attempt was by Korbinian Brodmann, published in German in 1909 as “Comparative Localization Studies in the Brain Cortex, its Fundamentals Represented on the Basis of its Cellular Architecture.” He divided the cerebral cortex (the outermost “layer” of the brain) into 52 regions based on the structure of cells (cytoarchitecture) sampled from different sections of brains taken from 64 different mammalian species, including humans (see Figure 1). Although Brodmann’s studies were purely anatomical, he wrote: “my ultimate goal was the advancement of a theory of function and its pathological deviations.” Nevertheless, he rejected what he saw as naive attempts at functional localization:
[Dressing] up the individual layers with terms borrowed from physiology or psychology…and all similar expressions that one encounters repeatedly today, especially in the psychiatric and neurological literature, are utterly devoid of any factual basis; they are purely arbitrary fictions and only destined to cause confusion in uncertain minds.
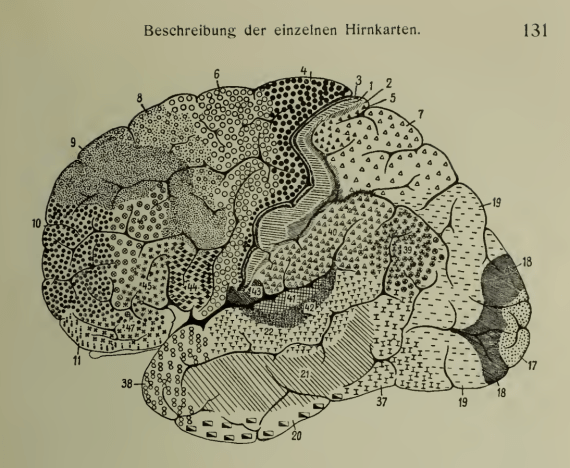
Over a century later, many researchers continue to refer to “Brodmann’s area” numbers as general orientation markers. More recently (see Figure 2), using data from the Human Connectome Project and supervised machine learning techniques, a team of researchers characterized 180 areas in each hemisphere — 97 new areas and 83 areas identified in previous work (Glasser et al. 2016). This study used a “multi-modal” technique which included cytoarchitecture, like Brodmann, but also connectivity, topography and function. For the latter, the study used data from “task functional MRI (tfMRI) contrasts,” wherein resting state measures are compared with measures taken during seven different tasks.

One of these tasks was language processing using a procedure developed by Binder et al. (2011) wherein participants read a short fable and then are asked a forced-choice question. Glasser et al. found reasonable evidence associating this language task with previously identified components of the “language network” (for recent overviews of the quest to localize the language network, see Frederici 2017 and Fitch 2018, both largely within the generative tradition). Specifically, these are Broca’s area (roughly 44) and Wernicke’s area (roughly PSL), and also identified an additional area, which they call 55b). Their findings also agreed with previous work going back to Broca on the “left-lateralization” of the language network—which means not that language is only in the left hemisphere (as some folk theories purport), but simply the left areas show more activity in response to the language task than in homologous areas in the right hemisphere (an early finding which inspired Jaynes’ Bicameral Mind hypothesis)
Does this mean we have discovered the “language module” theorized by Fodor, Chomsky, and others? Not quite, for three reasons. First, Glasser et al. found if they removed the functional task data, their classifier was nearly as accurate at identifying parcels. Second, the parcels were averaged over a couple hundred brains, and yet the classifier was still able to identify parcels in atypical brains (whether this translated into changes in functionality was outside the scope of the study).
Third, and most important for our purposes, this work does not—and the researchers do not attempt to—determine whether parcels are uniquely specialized (or encapsulated in Fodor’s terms). That is, while we can roughly identify a language network implicating relatively consistent areas across different brains, this does not demonstrate that such structures are necessary and sufficient for human language, and solely used for this purpose. Indeed, language may be a “repurposing” brain parcels used for (evolutionarily or developmentally older) processes. This is precisely the thesis of neural “exaption.”
What is Exaption?
In the last few decades several new frameworks—under labels like neural reuse, neuronal recycling, neural exploitation, or massive redeployment—attempt to offer a bridge between the modularity assumptions which undergird most neuroanatomical research, on one hand, and the connectionist assumptions which spurred advancements in artificial intelligence research and anthropology on the other. Such frameworks also attempt to account for the fact there is some consistency in activation across individuals, which does look a little bit like modularity.
The basic idea is exaption (also called exaptation): some biological tendencies or anatomical constraints may predispose certain areas of the brain to be implicated in certain cognitive functions, but these same areas may be recycled, repurposed, or reused for other functions. Exemplars of this approach are Stanislas Dehaene’s Reading in the Brain and Michael Anderson’s After Phrenology.
Perhaps the easiest way to give a sense of what this entails is to consider cases of neurodiversity, specifically the anthropologist Greg Downey’s essay on the use of echolocation by the visually impaired. While folk understandings may suggest that hearing becomes “better” in those with limited sight, this is not quite the case. Rather, one study finds, when listening to “ a recording [which] had echoes, parts of the brain associated with visual perception in sighted individuals became extremely active.” In other words, the brain repurposed the visual cortex as a result of the individual’s practices. While most humans have limited echolocation abilities and the potential to develop this skill, only some will put in the requisite practice.
Another strand of research supporting neural exaption falls under the heading of “conceptual metaphor theory” (itself a subfield of cognitive linguistics). The basic argument from this literature is that people tend to reason about (target) domains they have had little direct experience with by analogy to (source) domains with which they have had much direct experience (e.g. the nation is a family). As argued in Lakoff and Johnson’s famous Metaphors We Live By, this metaphorical mapping is not just figurative or linguistic, but rather a pre-linguistic conceptual mappings, and an—if not the—essential part of all cognition (Hofstadter and Sander 2013). Therefore, thinking or talking about even very abstract concepts re-activates a coalition of neural associations, many of which are fundamentally adapted to the mundane sensorimotor task of navigating our bodies through space. As we discuss in our forthcoming paper, “Schemas and Frames” (Wood et al. 2018), because talking and thinking recruit areas of our neural system often deployed in other activities—and at time-scales faster than conscious awareness can adequately attend—our biography of embodiment channels our reasoning in ways that seem intuitive and yet are constrained by the pragmatic patterns of those source domains. This is fully compatible with the dispositional theory of the mental Omar discusses.
What does this mean for sociology? I think there are numerous implications and we are just beginning to see how generative these insights are for our field. Here, I will limit myself to discussing just two, specifically related to how we tend to think about the role of language in our work. First, for an actor, knowing what text or talk means involves an actual embodied simulation of the practices it implies, very often (but not necessarily) in service of those practices in the moment (Binder and Desai 2011). Therefore, language should not be understood as an autonomous realm wherein meanings are produced by the internal interplay of contrastive differences within an always deferred linguistic system. Rather, following the later Wittgenstein in the Philosophical Investigations, “in most cases, the meaning of a word is its use.” Furthermore, as our embodiment is largely (but certainly not completely) shared across very different peoples (for example, most of us experience gravity all the time), there is a significant amount of shared semantics across diverse peoples (Wierzbicka 1996)—indeed without this, translation would likely be impossible.
Second, the repurposing of vocabulary commonly used in one context into a new context will often involve the analogical transfer of traces of the old context. This is because invoking such language activates a simulation of practices from the old context while one is in the new context. (Although this is dependent upon the accrued biographies of the individuals involved). This suggests that our language can be constraining in predictable ways, but not because the language itself has a structure or code rendering certain possibilities unthinkable. Rather, it is that language is the manifestation of a habit inextricably involved in a cascade of other habits, making it easier to execute (and therefore more probable for) some actions or thoughts over others. For example, as Barry Schwartz argued in his (criminally under-appreciated) Vertical Classification, it is nearly universal that UP is associated with power and also the morally good as a result of (near-universal) practices we encounter as babies and children. This helps explain the persistence of the “height premium” in the labor market (e.g., Lundborg, Nystedt, and Rooth 2014).
References
Binder, Jeffrey R. et al. 2011. “Mapping Anterior Temporal Lobe Language Areas with fMRI: A Multicenter Normative Study.” NeuroImage 54(2):1465–75.
Binder, Jeffrey R. and Rutvik H. Desai. 2011. “The Neurobiology of Semantic Memory.” Trends in Cognitive Sciences 15(11):527–36.
Dronkers, N. F. 2000. “The Pursuit of Brain–language Relationships.” Brain and Language. Retrieved (http://www.ebire.org/aphasia/dronkers/the_pursuit.pdf).
Fitch, W. Tecumseh. 2018. “The Biology and Evolution of Speech: A Comparative Analysis.” Annual Review of Linguistics 4(1):255–79.
Friederici, Angela D. 2017. Language in Our Brain: The Origins of a Uniquely Human Capacity. MIT Press.
Glasser, Matthew F. et al. 2016. “A Multi-Modal Parcellation of Human Cerebral Cortex.” Nature 536(7615):171–78.
Grodzinsky, Yosef and Katrin Amunts. 2006. Broca’s Region. Oxford University Press, USA.
Hofstadter, Douglas and Emmanuel Sander. 2013. Surfaces and Essences: Analogy as the Fuel and Fire of Thinking. Basic Books.
Lindenberg, Robert, Heiner Fangerau, and Rüdiger J. Seitz. 2007. “‘Broca’s Area’ as a Collective Term?” Brain and Language 102(1):22–29.
Lundborg, Petter, Paul Nystedt, and Dan-Olof Rooth. 2014. “Height and Earnings: The Role of Cognitive and Noncognitive Skills.” The Journal of Human Resources 49(1):141–66.
Viola, Marco and Elia Zanin. 2017. “The Standard Ontological Framework of Cognitive Neuroscience: Some Lessons from Broca’s Area.” Philosophical Psychology 30(7):945–69.
Wierzbicka, Anna. 1996. Semantics: Primes and Universals. Oxford University Press, UK.
Wood, Michael Lee, Dustin S. Stoltz, Justin Van Ness, and Marshall A. Taylor. 2018. “Schemas and Frames.” Retrieved (https://osf.io/preprints/socarxiv/b3u48/).
One thought on “Exaption: Alternatives to the Modular Brain, Part II14 min read”